- SÉLECTION NATURELLE (biologie)
- SÉLECTION NATURELLE (biologie)La notion de sélection (du latin selectio , choix) revêt une importance capitale en biologie, qu’elle soit appliquée ou fondamentale. Le tri qu’implique toute sélection porte ici sur la diversité phénotypique et génotypique des organismes, diversité dont l’existence a été reconnue, au sein de toute population, et a fortiori de toute espèce. La sélection dirigée par l’homme est qualifiée d’artificielle, par opposition à la sélection naturelle, force évolutive d’une puissance fantastique, dont le moteur est la nature même, jouant par l’interaction de tous ses paramètres depuis l’origine de la vie.Sélection artificielleC’est par la mise en œuvre de méthodes sélectives que l’homme, depuis des temps reculés, a réussi à améliorer des variétés de plantes utiles: légumes, céréales, arbres fruitiers et des races d’animaux domestiques, ou même à créer de nouvelles variétés ou races.Les recherches orientées des sélectionneurs et des agronomes modernes, avertis des données de la génétique formelle et quantitative, n’ont que de lointains rapports avec les essais désordonnés des agriculteurs ou éleveurs des siècles passés, dont les succès pragmatiques ne sauraient pourtant être minimisés. Les «nouveautés» ont souvent pour origine une simple mutation ou une nouvelle combinaison génique, conférant à un individu donné un caractère inhabituel, plus ou moins frappant, que les hommes, par intérêt ou curiosité, se sont efforcés de perpétuer. Ainsi sont nées d’innombrables races stabilisées, parfois belles, ou étranges, ou monstrueuses: races de chiens, de chats, de volailles, moutons et bœufs sans cornes, chiens et bovins bouledogues, végétaux nains ou pleureurs, fleurs doubles, variétés de pelage ou de couleur n’en sont que quelques exemples. L’obtention de races pures est plus ou moins aisée: la sélection d’un gène récessif est beaucoup plus rapide que celle d’un allèle dominant (les hétérozygotes étant le plus souvent indiscernables des homozygotes dominants). Le danger que représentent pour le maintien de telles races des croisements successifs entre descendants (in-breeding ), donc une consanguinité trop poussée, ne doit jamais être sous-estimé.Le succès de la sélection de caractères qualitatifs ne saurait transgresser le stade de génotype homozygote. Il en va de même pour les caractères quantitatifs dont l’estimation nécessite des mesures. Sur le haricot, végétal se reproduisant par autofécondation, W. L. Johansen (1903) montra que la sélection devenait très vite inefficace, chaque graine d’un lot commercial correspondant à un génotype homozygote; les différences de poids observées dans des lots de graines de même génotype expriment simplement une inéluctable variation phénotypique due à l’action du milieu.Au contraire, chez les organismes se reproduisant par fécondation croisée (cas de nombreux végétaux et de la grande majorité des animaux), la sélection des caractères quantitatifs, dont le résultat dépend naturellement du choix des reproducteurs, peut être poursuivie dans une même direction et demeurer efficace bien au-delà de la première génération, jusqu’à une limite qui ne sera que très graduellement atteinte. Ce fait tient à ce que les caractères quantitatifs sont contrôlés par de nombreux gènes dont les effets majorants ou minorants sont additifs. L’accumulation progressive de gènes «fastes» a permis des performances agronomiques d’un intérêt primordial: par exemple, la betterave sucrière, qui, au début du XIXe siècle, renfermait 6 p. 100 de saccharose, atteint aujourd’hui en Europe une teneur de 18 à 20 p. 100. Une sélection prolongée peut conduire à de véritables hyperthélies: les coqs Phénix ont des faucilles atteignant cinq à six mètres de longueur.On conçoit que le succès d’une sélection dépende de la richesse du stock génique initial; il est donc favorable de procéder à partir non seulement d’individus suffisamment nombreux, mais aussi de descendants de croisements entre souches ou races différentes. Il est remarquable qu’une telle méthode ait été employée, grâce à une sorte de bon sens prémonitoire, bien avant la découverte des lois de l’hérédité, comme en témoigne l’exemple des porcs.Peu différente des sangliers à son origine (fin du XVIIIe s.), la race de porc Berkshire fut améliorée après de nombreux croisements avec diverses races (cochon chinois, sanglier demi-sauvage, cochon napolitain, porc Essex-Suffolk) pour donner finalement (vers 1900) la race actuelle sélectionnée pour sa viande.L’homme a été tenté, depuis l’Antiquité grecque, par l’idée d’améliorer les populations de sa propre espèce. En théorie, un tel eugénisme ne pourrait avoir que d’heureuses conséquences sur la vie sociale, par suppression des tares. Mais si l’on peut envisager un succès biologique en ce qui concerne les tares dominantes (à condition d’appliquer à l’homme les techniques zootechniques d’extermination ou de castration des «anormaux»), une élimination des tares récessives, de loin les plus nombreuses et les plus délétères, s’avère rigoureusement impossible. Seule une entrave aux mariages consanguins peut réduire la fréquence des maladies récessives [cf. EUGÉNIQUE ET EUGÉNISME].Si dans l’ensemble, les résultats de la sélection artificielle sont et continuent d’être remarquables en ce qui concerne la biologie appliquée, ils demeurent, d’un point de vue évolutif, limités par le facteur temps (quelques milliers d’années seulement) à la différenciation de simples races. Ils ont cependant fourni à Darwin (après que le fait d’évolution se fut imposé à lui au cours du voyage sur le Beagle, et qu’il eut eu connaissance de l’œuvre de Malthus) et à A. R. Wallace le modèle élémentaire de cette force omnipotente depuis quelques trois milliards d’années qu’est la sélection naturelle.Sélection naturelleLes diverses conceptions«Étant donné que plus d’individus sont produits qu’il n’en peut survivre, il doit exister dans chaque cas une lutte pour l’existence, soit entre un individu et un autre individu de la même espèce, soit entre individus d’espèces différentes. Peut-on, dès lors, considérer comme improbable, puisque des variations utiles à l’homme sont manifestement survenues, que d’autres variations utiles en quelque manière à chaque être vivant, dans la grande et complexe bataille pour la vie, se soient parfois produites au cours de milliers de générations? S’il en va ainsi, pouvons-nous douter (en nous rappelant que bien plus d’individus naissent qu’il n’en peut survivre) que les individus possédant un avantage quelconque, si minime soit-il, sur les autres auraient une meilleure chance de survivre et de procréer leur propre type? Inversement, nous pouvons être assurés que toute variation, délétère à quelque degré, serait impitoyablement éliminée. Cette préservation des variations favorables et ce rejet des variations défavorables, je l’appelle la sélection naturelle.»C’est ainsi que Charles Darwin, dans The Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the struggle for Life (1859), définissait la sélection naturelle sur laquelle reposait sa théorie de l’évolution.Des présélectionnistes avaient eu, avant Darwin et Wallace, l’idée d’un jeu sélectif, mais ils n’attribuaient à la sélection qu’un rôle conservateur, normalisateur, d’ailleurs réel et d’une extrême importance. On doit à Darwin d’avoir compris le pouvoir novateur de la sélection naturelle, processus responsable de la formation des espèces et du progrès continu des adaptations. La conception darwinienne reposait sur l’existence, au sein de toute population, de variations phénotypiques (pour une part transmissibles héréditairement), dont certaines s’avèrent plus favorisées que d’autres; les catégories qui réalisent les meilleures adaptations au milieu laissent, en moyenne, plus de descendants que les autres, ce qui entraîne une évolution adaptative des populations.L’argumentation de Darwin, si brillante et féconde qu’elle fût, resta pourtant entachée d’une lacune essentielle: Darwin ignora jusqu’à sa mort (1882) l’œuvre de J. Mendel (1865). La redécouverte des lois de l’hérédité (1900) aurait dû peser immédiatement dans les controverses qui opposaient partisans et détracteurs du darwinisme; il n’en fut rien, au point que, vers 1920, et notamment en France, la plupart des biologistes s’abandonnaient à un scepticisme plus ou moins vitaliste et se désintéressaient pratiquement d’un problème tenu pour insoluble.Les recherches entreprises indépendamment, notamment par R. A. Fisher (1930), S. Wright (1931), J. B. S. Haldane (1932), G. Teissier et P. L’Héritier (1933), devaient heureusement assurer le regain du darwinisme en le faisant bénéficier de l’acquis génétique et conduire, par intégration des données accumulées par les systématiciens de populations et les écologistes, à la théorie synthétique de l’évolution. Leur mérite essentiel a été de faire comprendre que la sélection naturelle était un processus statistique , jouant en termes de probabilités sur des populations naturelles; il n’est pas question d’une survivance automatique et exclusive du plus favorisé, de la fameuse «survivance du plus apte», selon le slogan de Spencer, mais de tenir compte du simple fait que le plus apte a, dans des conditions définies, une probabilité plus grande de transmettre ses caractères. La sélection naturelle assure en fait la transmission statistiquement préférentielle de ces génotypes (E. Mayr, 1964).On comprend, par là même, que la sélection sexuelle soit considérée par beaucoup d’auteurs comme une forme particulière de sélection naturelle. Elle consiste dans le fait que les mâles les plus vigoureux (dans les contacts qui préludent aux accouplements), ou les plus entreprenants, ou les plus subtils, prennent le pas sur leurs rivaux dans la séduction et la possession des femelles, trop souvent considérées, à tort, comme passives. Elle serait responsable, pour beaucoup, des manifestations le plus souvent étonnantes du dimorphisme sexuel chez la plupart des espèces gonochoriques, manifestations morphologiques (développement de structures hyperthéliques chez les mâles, couleurs éclatantes de l’un des deux sexes, au moins au moment des pariades) ou éthologiques (postures, chants, danses, offrandes). L’importance relative d’une vigueur liée à l’hétérozygotie des mâles et de facteurs comportementaux fait aujourd’hui l’objet de nombreuses recherches expérimentales d’un grand intérêt.Valeur sélective des génotypesUne population vivant dans un milieu fini, ne disposant que d’une quantité de nourriture limitée, se maintient théoriquement à un effectif globalement stable d’une génération à l’autre (avec, en réalité, des fluctuations parfois importantes, dues aux variations climatiques). Un couple ne laisse en moyenne, dans le cas d’une reproduction sexuée biparentale, que deux descendants, bien que le potentiel de fécondité physiologique soit très supérieur. Ce fait évident permet d’évoquer l’une des critiques qui ont été le plus souvent formulées contre la sélection naturelle; s’il ne subsiste que deux adultes sur une ponte de 600 000 œufs, est-il si clair que ce seront les plus aptes qui survivront? Il est vrai que l’élimination massale des gamètes, des œufs ou des larves est largement aléatoire, mais on ne saurait pour autant écarter l’idée d’une destruction différentielle: une larve planctonique dotée d’un meilleur mécanisme de sustentation est nécessairement favorisée statistiquement par rapport aux autres. C’est d’ailleurs sur l’échantillon des survivants parvenus à maturité sexuelle qu’interviennent, à côté de facteurs de chance ou de malchance, des forces sélectives intrinsèques. D’un couple à l’autre, le nombre de descendants varie beaucoup et dépend, abstraction faite de facteurs fortuits, de la structure génétique des zygotes. Or, on sait aujourd’hui qu’il existe, dans toute population naturelle, un éventail plus ou moins étendu de phénotypes, correspondant à une gamme génotypique encore plus large. Ces génotypes possèdent des caractéristiques physiologiques différentes, affectant leur vitalité et leur fécondité. «On peut alors concevoir que chacun des génotypes, présents même à un moment ou à un autre dans une population, puisse être caractérisé par un taux de survie et par un taux de fécondité 猪 particuliers. La valeur sélective 靖 du génotype est, par définition, le produit de ces deux nombres: 靖 =猪; elle est l’équivalent, pour l’ensemble des représentants d’un génotype, du taux net de reproduction que définissent les démographes pour l’ensemble d’une population, et est égale au nombre moyen de descendants laissés par un individu appartenant au phénotype considéré» (Teissier, 1955).L’introduction de la notion de valeur sélective d’un génotype dans les calculs de génétique des populations, discipline dont le but original était d’apprécier objectivement le rôle attribué à la sélection naturelle, fournit le modèle élémentaire d’un processus évolutif. En effet, si l’on considère, dans une population d’effectif infini, un couple d’allèles Aa , si l’on suppose qu’il n’y a pas de mutation ni de choix des partenaires et que les trois génotypes AA , Aa et aa sont également viables et féconds, on démontre aisément que les fréquences respectives p de A et q de a (p + q = 1) restent constantes (loi de Hardy-Weinberg), les fréquences des génotypes étant:
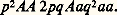 Cette expression qui établirait, si les préalables étaient respectés, un fixisme absolu de la population devient, après une génération, en sélection zygotique et pour des gènes autosomiques, en attribuant respectivement aux génotypes AA , Aa et aa les valeurs sélectives 靖1, 靖2, et 靖3: 靖1p 2AA , 2 靖2pqAa , 靖3q 2aa ; on en déduit facilement les variations p et q de la fréquence des allèles A et a , étant entendu qu’interviennent dans les formules les rapports entre les trois vigueurs sélectives et non les valeurs sélectives absolues.Le signe de p et q détermine évidemment le sens dans lequel évoluera la population. Si la sélection s’effectue à coefficients constants, il ne peut s’installer d’équilibre stable entre les allèles A et a que si 靖2 est à la fois supérieur à 靖1 et à 靖3. En dehors de cette condition nécessaire et suffisante, la sélection éliminera l’un des allèles. D’une manière générale, la sélection tend à faire disparaître tous les allèles d’un gène autres que le normal, cette élimination étant plus ou moins rapide suivant le caractère plus ou moins délétère des génotypes; elle n’est en tous cas pas instantanée, et un allèle désavantageux peut subsister indéfiniment dans une population s’il y réapparaît constamment par mutation, sa fréquence dépendant du degré de dominance.Le problème est beaucoup plus complexe si la sélection s’effectue à coefficients variables, c’est-à-dire lorsque les valeurs sélectives des génotypes, pour un locus donné, dépendent de la composition même de la population. De telles situations, réalisées chaque fois que les génotypes en présence n’ont pas exactement les mêmes besoins qualitatifs ou nécessitent des niches écologiques différentes, ont été analysées pour la première fois par G. Teissier (1942); C. Petit en a nettement montré l’importance dans le maintien des gènes rares.Si, dans les sélections à coefficients constants, la valeur sélective moyenne 諸, exprimée par la somme des produits: fréquence d’un génotype 憐 valeur sélective de ce génotype, tend constamment à s’accroître, elle peut au contraire décroître dans certains types de sélection à coefficients variables, par exemple dans le cas de compétition entre différents membres d’une population, la marche de la sélection conduisant alors à l’extinction de la population.Si l’on considère le cas d’un unique couple d’allèles pour permettre l’approche mathématique des problèmes de la sélection naturelle, c’est au niveau des organismes, compte tenu de leurs caractères phénotypiques, qu’opère la sélection naturelle. Sachant qu’une drosophile est le résultat de l’interaction d’environ 100 000 gènes, que chaque Vertébré supérieur exprime l’interaction d’un nombre de gènes probablement plus grand encore, que la valeur sélective d’un individu est un compromis entre des forces sélectives alléliques antagonistes, que les valeurs sélectives peuvent varier considérablement suivant les étapes d’un cycle biologique, on conçoit déjà les difficultés surhumaines que soulève l’appréciation réelle de la valeur sélective globale d’un génotype donné. À cela s’ajoute le fait, non moins important, que la valeur sélective d’un génotype n’est pas une propriété intrinsèque de ce dernier, mais dépend très largement des conditions perpétuellement changeantes (toutefois, en général, à l’intérieur de limites assez étroites) du milieu naturel dans lequel est placée la population à laquelle appartient l’individu considéré, soumise à des compétitions interspécifiques et notamment aux attaques de prédateurs ou de parasites.L’estimation précise de la valeur sélective d’un génotype apparaît ainsi, eu égard aux possibilités humaines, comme presque aussi inaccessible que les phénomènes évolutifs qu’elle dirige; cela ne diminue en rien l’efficacité d’une force dont la présence a été vérifiée expérimentalement, dans des conditions nécessairement fort simples par rapport à celles qui règnent dans le plus simple des milieux naturels.Sélection et évolutionL’étude expérimentale de la sélection a été abordée par L’Héritier et Teissier (1933), grâce à l’emploi des cages à population de drosophiles ou démomètres, cages dans lesquelles la population atteint rapidement un effectif important et stable et où les mouches, maintenues à une température et à un degré hygrométrique définis, disposent d’une nourriture plus ou moins riche, renouvelée à un rythme régulier. La compétition entre drosophiles sauvages et mutantes a établi la validité des conclusions théoriques de la génétique des populations; certains gènes sont éliminés, alors que, pour d’autres, il s’établit entre eux et l’allèle sauvage un état d’équilibre qui peut résulter soit d’une supériorité des hétérozygotes, soit du jeu d’une concurrence larvaire (sélection à coefficients variables). La même technique des démomètres a été utilisée ultérieurement par de nombreux auteurs et notamment par T. Dobzhansky et ses collaborateurs pour l’analyse du maintien d’un polymorphisme chromosomique: l’intérêt majeur de ces dernières observations, dont l’interprétation est facilitée par une suprématie des hétérozygotes, a été d’établir, pour certaines inversions de Drosophila pseudo-obscura , un parallélisme entre les équilibres obtenus expérimentalement à diverses températures et les variations de fréquences saisonnières observées dans les populations naturelles. Le polymorphisme génétique de populations naturelles a fait d’ailleurs l’objet de nombreuses recherches récentes, notamment chez les Cepaea et les Sphéromes; la complexité des situations rencontrées explique que l’interprétation des résultats obtenus continue de susciter des controverses et nous demeure largement interdite.Le problème d’une comparaison entre la valeur adaptative des génotypes et celle de la population à laquelle ils appartiennent et qui représente l’unité évolutive réelle soulève lui aussi beaucoup de difficultés, notamment dans le choix d’une mesure permettant de quantifier l’adaptation d’une population à son milieu.L’évolution des fréquences géniques au sein d’une population, la meilleure adaptation des génotypes et des populations aux conditions du milieu sont autant de preuves immédiatement perceptibles de l’action permanente de la sélection naturelle. L’une des fonctions de celle-ci consiste dans l’élimination d’un excès de variabilité qui aurait fait du monde vivant, si la sélection n’avait pas joué, un véritable chaos. En continuant à chaque génération d’éliminer certains gènes et d’en promouvoir d’autres, elle assure une sorte de normalisation des populations et des espèces, dans la mesure où les conditions de l’environnement ne subissent pas d’altérations trop marquées. L’importance de cette sélection conservatrice a été perçue depuis longtemps, et encore soulignée par S. Ohno (1970). Mais, si les conditions de milieu subissent des modifications importantes, ce qui est bien souvent survenu à la surface du globe, la sélection naturelle interviendra, en effectuant de nouveaux tris entre les possibilités génétiques ou en imposant de nouvelles recombinaisons géniques, et modulera ainsi peu à peu les espèces nouvelles et, par la succession ininterrompue des générations, les catégories systématiques supérieures à l’espèce. Cette sélection novatrice apparaît ainsi comme l’un des moteurs essentiels, responsables de la formidable diversité, pourtant harmonieusement orientée, des règnes vivants.Est-ce à dire que la sélection naturelle explique à elle seule toute l’évolution? Disons tout d’abord qu’elle n’aurait jamais existé si une diversité génique ne s’était pas établie, probablement dès les premières manifestations de la vie, à la surface du globe, lui fournissant les éléments d’un choix: c’est dans ce matériel exploitable, d’une richesse difficilement imaginable, et constitué par les allèles et les recombinaisons, qu’elle a puisé, pour réaliser la diversité des espèces, les exemples de mimétisme qui nous stupéfient ou les souches d’insectes résistant en un temps record aux pesticides et celles de micro-organismes résistant aux antibiotiques. De plus, dans des populations de faibles effectifs, la fréquence des gènes peut subir une dérive, et l’on ne saurait minimiser l’effet Wright, susceptible d’intervenir à tout moment, par exemple lors d’une épidémie catastrophique ou de la fondation d’une nouvelle colonie par un petit nombre d’individus migrants, et qui, du moins temporairement, peut aller à l’encontre des probabilités sélectionnistes.De toute manière, dans l’interaction complexe des trois forces évolutives: pression de mutation, dérive génique et pression de sélection, cette dernière possède des atouts majeurs. Pour être longtemps demeurée occulte, à force d’avoir agi partout et constamment, la pression de sélection n’en a pas moins modelé le monde vivant.
Cette expression qui établirait, si les préalables étaient respectés, un fixisme absolu de la population devient, après une génération, en sélection zygotique et pour des gènes autosomiques, en attribuant respectivement aux génotypes AA , Aa et aa les valeurs sélectives 靖1, 靖2, et 靖3: 靖1p 2AA , 2 靖2pqAa , 靖3q 2aa ; on en déduit facilement les variations p et q de la fréquence des allèles A et a , étant entendu qu’interviennent dans les formules les rapports entre les trois vigueurs sélectives et non les valeurs sélectives absolues.Le signe de p et q détermine évidemment le sens dans lequel évoluera la population. Si la sélection s’effectue à coefficients constants, il ne peut s’installer d’équilibre stable entre les allèles A et a que si 靖2 est à la fois supérieur à 靖1 et à 靖3. En dehors de cette condition nécessaire et suffisante, la sélection éliminera l’un des allèles. D’une manière générale, la sélection tend à faire disparaître tous les allèles d’un gène autres que le normal, cette élimination étant plus ou moins rapide suivant le caractère plus ou moins délétère des génotypes; elle n’est en tous cas pas instantanée, et un allèle désavantageux peut subsister indéfiniment dans une population s’il y réapparaît constamment par mutation, sa fréquence dépendant du degré de dominance.Le problème est beaucoup plus complexe si la sélection s’effectue à coefficients variables, c’est-à-dire lorsque les valeurs sélectives des génotypes, pour un locus donné, dépendent de la composition même de la population. De telles situations, réalisées chaque fois que les génotypes en présence n’ont pas exactement les mêmes besoins qualitatifs ou nécessitent des niches écologiques différentes, ont été analysées pour la première fois par G. Teissier (1942); C. Petit en a nettement montré l’importance dans le maintien des gènes rares.Si, dans les sélections à coefficients constants, la valeur sélective moyenne 諸, exprimée par la somme des produits: fréquence d’un génotype 憐 valeur sélective de ce génotype, tend constamment à s’accroître, elle peut au contraire décroître dans certains types de sélection à coefficients variables, par exemple dans le cas de compétition entre différents membres d’une population, la marche de la sélection conduisant alors à l’extinction de la population.Si l’on considère le cas d’un unique couple d’allèles pour permettre l’approche mathématique des problèmes de la sélection naturelle, c’est au niveau des organismes, compte tenu de leurs caractères phénotypiques, qu’opère la sélection naturelle. Sachant qu’une drosophile est le résultat de l’interaction d’environ 100 000 gènes, que chaque Vertébré supérieur exprime l’interaction d’un nombre de gènes probablement plus grand encore, que la valeur sélective d’un individu est un compromis entre des forces sélectives alléliques antagonistes, que les valeurs sélectives peuvent varier considérablement suivant les étapes d’un cycle biologique, on conçoit déjà les difficultés surhumaines que soulève l’appréciation réelle de la valeur sélective globale d’un génotype donné. À cela s’ajoute le fait, non moins important, que la valeur sélective d’un génotype n’est pas une propriété intrinsèque de ce dernier, mais dépend très largement des conditions perpétuellement changeantes (toutefois, en général, à l’intérieur de limites assez étroites) du milieu naturel dans lequel est placée la population à laquelle appartient l’individu considéré, soumise à des compétitions interspécifiques et notamment aux attaques de prédateurs ou de parasites.L’estimation précise de la valeur sélective d’un génotype apparaît ainsi, eu égard aux possibilités humaines, comme presque aussi inaccessible que les phénomènes évolutifs qu’elle dirige; cela ne diminue en rien l’efficacité d’une force dont la présence a été vérifiée expérimentalement, dans des conditions nécessairement fort simples par rapport à celles qui règnent dans le plus simple des milieux naturels.Sélection et évolutionL’étude expérimentale de la sélection a été abordée par L’Héritier et Teissier (1933), grâce à l’emploi des cages à population de drosophiles ou démomètres, cages dans lesquelles la population atteint rapidement un effectif important et stable et où les mouches, maintenues à une température et à un degré hygrométrique définis, disposent d’une nourriture plus ou moins riche, renouvelée à un rythme régulier. La compétition entre drosophiles sauvages et mutantes a établi la validité des conclusions théoriques de la génétique des populations; certains gènes sont éliminés, alors que, pour d’autres, il s’établit entre eux et l’allèle sauvage un état d’équilibre qui peut résulter soit d’une supériorité des hétérozygotes, soit du jeu d’une concurrence larvaire (sélection à coefficients variables). La même technique des démomètres a été utilisée ultérieurement par de nombreux auteurs et notamment par T. Dobzhansky et ses collaborateurs pour l’analyse du maintien d’un polymorphisme chromosomique: l’intérêt majeur de ces dernières observations, dont l’interprétation est facilitée par une suprématie des hétérozygotes, a été d’établir, pour certaines inversions de Drosophila pseudo-obscura , un parallélisme entre les équilibres obtenus expérimentalement à diverses températures et les variations de fréquences saisonnières observées dans les populations naturelles. Le polymorphisme génétique de populations naturelles a fait d’ailleurs l’objet de nombreuses recherches récentes, notamment chez les Cepaea et les Sphéromes; la complexité des situations rencontrées explique que l’interprétation des résultats obtenus continue de susciter des controverses et nous demeure largement interdite.Le problème d’une comparaison entre la valeur adaptative des génotypes et celle de la population à laquelle ils appartiennent et qui représente l’unité évolutive réelle soulève lui aussi beaucoup de difficultés, notamment dans le choix d’une mesure permettant de quantifier l’adaptation d’une population à son milieu.L’évolution des fréquences géniques au sein d’une population, la meilleure adaptation des génotypes et des populations aux conditions du milieu sont autant de preuves immédiatement perceptibles de l’action permanente de la sélection naturelle. L’une des fonctions de celle-ci consiste dans l’élimination d’un excès de variabilité qui aurait fait du monde vivant, si la sélection n’avait pas joué, un véritable chaos. En continuant à chaque génération d’éliminer certains gènes et d’en promouvoir d’autres, elle assure une sorte de normalisation des populations et des espèces, dans la mesure où les conditions de l’environnement ne subissent pas d’altérations trop marquées. L’importance de cette sélection conservatrice a été perçue depuis longtemps, et encore soulignée par S. Ohno (1970). Mais, si les conditions de milieu subissent des modifications importantes, ce qui est bien souvent survenu à la surface du globe, la sélection naturelle interviendra, en effectuant de nouveaux tris entre les possibilités génétiques ou en imposant de nouvelles recombinaisons géniques, et modulera ainsi peu à peu les espèces nouvelles et, par la succession ininterrompue des générations, les catégories systématiques supérieures à l’espèce. Cette sélection novatrice apparaît ainsi comme l’un des moteurs essentiels, responsables de la formidable diversité, pourtant harmonieusement orientée, des règnes vivants.Est-ce à dire que la sélection naturelle explique à elle seule toute l’évolution? Disons tout d’abord qu’elle n’aurait jamais existé si une diversité génique ne s’était pas établie, probablement dès les premières manifestations de la vie, à la surface du globe, lui fournissant les éléments d’un choix: c’est dans ce matériel exploitable, d’une richesse difficilement imaginable, et constitué par les allèles et les recombinaisons, qu’elle a puisé, pour réaliser la diversité des espèces, les exemples de mimétisme qui nous stupéfient ou les souches d’insectes résistant en un temps record aux pesticides et celles de micro-organismes résistant aux antibiotiques. De plus, dans des populations de faibles effectifs, la fréquence des gènes peut subir une dérive, et l’on ne saurait minimiser l’effet Wright, susceptible d’intervenir à tout moment, par exemple lors d’une épidémie catastrophique ou de la fondation d’une nouvelle colonie par un petit nombre d’individus migrants, et qui, du moins temporairement, peut aller à l’encontre des probabilités sélectionnistes.De toute manière, dans l’interaction complexe des trois forces évolutives: pression de mutation, dérive génique et pression de sélection, cette dernière possède des atouts majeurs. Pour être longtemps demeurée occulte, à force d’avoir agi partout et constamment, la pression de sélection n’en a pas moins modelé le monde vivant.
Encyclopédie Universelle. 2012.